Tiny Condensates, Mastering Synaptic Communication: Literature Interpretation of SNAP25 LLPS Mechanism
I. Research Background
Precise and rapid presynaptic neurotransmitter release is highly dependent on the SNARE complex-mediated synaptic exocytosis process. Three core SNARE proteins—Syntaxin-1, SNAP25, and VAMP2—bind together to form a four-helix trans-SNARE complex, bringing vesicles closer to the cell membrane and initiating the membrane fusion process. Structural abnormalities and gene mutations in SNAP25 directly disrupt synaptic signal transmission, and heterozygous SNAP25 mutations have been detected in a large number of patients with developmental epileptic encephalopathy (DEE). Previous microscopic imaging studies have observed that SNAP25 and Syntaxin-1 form nanoscale protein clusters on the synaptic membrane, and this aggregated state directly affects vesicle tethering, anchoring, and release initiation. However, for a long time, no study has explained the molecular driving force of protein clustering. Liquid-liquid phase separation (LLPS), as an assembly method for membrane-less compartments in cells, has been proven to regulate the spatial aggregation of synaptic scaffold proteins and synaptotagmin 1. Whether SNARE proteins, as the core of membrane fusion, rely on phase separation for local enrichment has remained a gap in the field. In 2026, the team of Min Zhu, Yinghui Liu, and Haijia Yu published a study titled SNAP25 undergoes phase separation to facilitate the synaptic vesicle fusion machinery in Cell Reports. Combining the unique molecular structure of SNAP25—a large intrinsically disordered linker region between the two SNARE motifs, with cysteines in the linker region that can undergo palmitoylation modification—and previous experimental conclusions that Syntaxin-1 enhances SNAP25 palmitoylation, the authors proposed a complete scientific hypothesis: SNAP25 undergoes liquid-liquid phase separation through its disordered linker region, with palmitoylation modification and Syntaxin-1 jointly regulating condensate formation. The droplets produced by phase separation can recruit all SNARE components to promote ternary complex assembly. DEE pathogenic mutations disrupt phase separation and block the membrane fusion pathway, thereby completely filling the research gap in the mechanism of SNARE protein aggregation.
II. Research Methods
This study integrates multiple experimental systems including in vitro biochemical reconstitution, live-cell optical detection, primary neuron models, and artificial membrane systems. Experimental materials include wild-type and series mutant SNAP25, Syntaxin-1 intracellular domain, and VAMP2 cytoplasmic domain proteins expressed and purified from prokaryotic systems. Various fluorescently labeled fusion proteins, photo-induced phase separation OptoDroplet plasmids, and targeted lentiviral interference vectors for SNAP25 and palmitoyltransferase ZDHHC17 were also constructed. In vitro experiments established a standardized phase separation reaction system, using fluorescence confocal and DIC differential interference imaging to observe droplet morphology, and sedimentation experiments, turbidity quantification, circular dichroism, and dibromomethane tryptophan quenching to characterize protein phase separation and conformational changes. At the cellular level, HeLa cell knockdown-recovery system was used to eliminate overexpression interference, FRAP fluorescence recovery after photobleaching experiments verified the liquid dynamic properties of condensates, and the OptoDroplet light-controlled system visually observed protein co-condensation behavior in live cells. In vitro membrane reconstitution included three artificial membranes: supported lipid bilayer (SLB), giant unilamellar vesicles (GUVs), and small unilamellar vesicles (SUVs), simulating cell membrane and synaptic vesicle structures to reproduce membrane surface protein aggregation and vesicle recruitment processes. Cell function experiments used rat primary hippocampal neurons, verifying protein expression levels through Western blotting and observing punctate protein aggregation in neurons via confocal imaging. For disease mechanism studies, two DEE-related mutant proteins (I67N, I192N) were constructed, and SDS-resistant functional SNARE complex production was detected in 293T cells. All experiments were performed with three or more biological replicates, and t-tests and one-way ANOVA were used for statistical difference determination.
III. Results Analysis
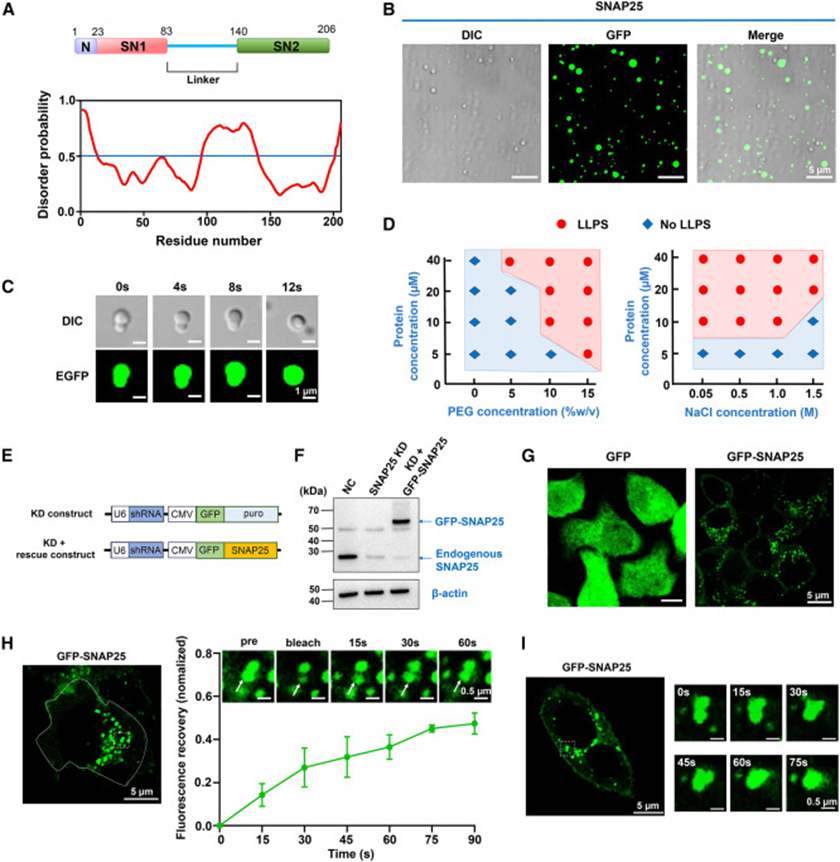
1. In vitro and live-cell experiments confirm SNAP25 possesses liquid-liquid phase separation properties
The study first analyzed the SNAP25 amino acid sequence using the protein disorder prediction tool PrDOS, revealing a large intrinsically disordered sequence in the linker region between the two SNARE motifs—a typical structural feature of phase-separating proteins. In the in vitro reconstitution system, purified EGFP-tagged SNAP25 spontaneously formed spherical droplets under physiological simulated conditions with the crowding agent PEG. Time-lapse imaging captured dynamic fusion of small droplets, intuitively demonstrating liquid properties. Gradient concentration and salt ion gradient experiments generated a complete phase diagram, proving that protein concentration, osmotic pressure, and molecular crowding collectively regulate phase separation, with physiological saline concentration range precisely satisfying condensate formation conditions. To exclude false aggregation caused by protein overexpression, the study constructed SNAP25 knockdown + endogenous level recovery HeLa cells. Control cells expressing free GFP showed uniform cytoplasmic fluorescence, while GFP-SNAP25 cells displayed numerous bright punctate condensates. FRAP bleaching experiments showed slow fluorescence recovery in condensates, reaching half recovery at 90 seconds. Time-lapse live-cell imaging also observed fusion of punctate condensates. Together, these cellular-level experiments confirm that SNAP25 can undergo liquid-phase condensation at physiological expression levels, with both in vitro and in vivo systems mutually validating SNAP25's intrinsic phase separation ability.
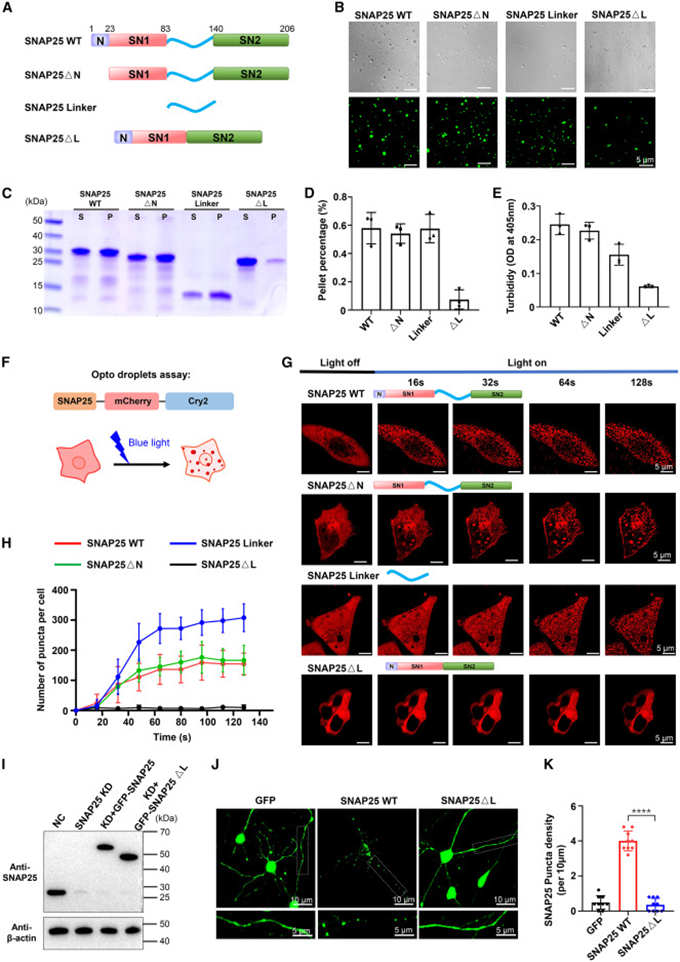
2. Disordered linker region is the core essential element for SNAP25 phase separation
The researchers designed a series of truncation mutants to dissect SNAP25 functional regions, removing the N-terminal peptide, completely deleting the middle linker region, and extracting the linker fragment alone for parallel in vitro phase separation experiments. Proteins with N-terminal deletion still formed regular spherical droplets, while linker-deleted mutants only produced amorphous flocculent precipitates. The isolated linker fragment formed even more droplets than the full-length protein. Sedimentation and turbidity quantification data further quantified these differences—the linker-deleted protein remained mostly in the supernatant, forming almost no precipitable condensates. The photo-induced OptoDroplet system simultaneously verified this pattern: under blue light stimulation, all proteins except the ΔL linker-deletion mutant formed intracellular condensation granules, with the isolated linker fragment producing significantly more granules. Finally, verification was repeated in rat primary hippocampal neurons. Wild-type SNAP25 formed clear fluorescent spots in neuronal soma and synaptic regions, while linker-deletion mutant protein showed completely uniform fluorescence diffusion. High-resolution imaging and hexanediol dissolution experiments confirmed that the punctate structures were typical liquid-phase condensates, fully demonstrating that the linker region is the sole core structure driving SNAP25 phase separation.
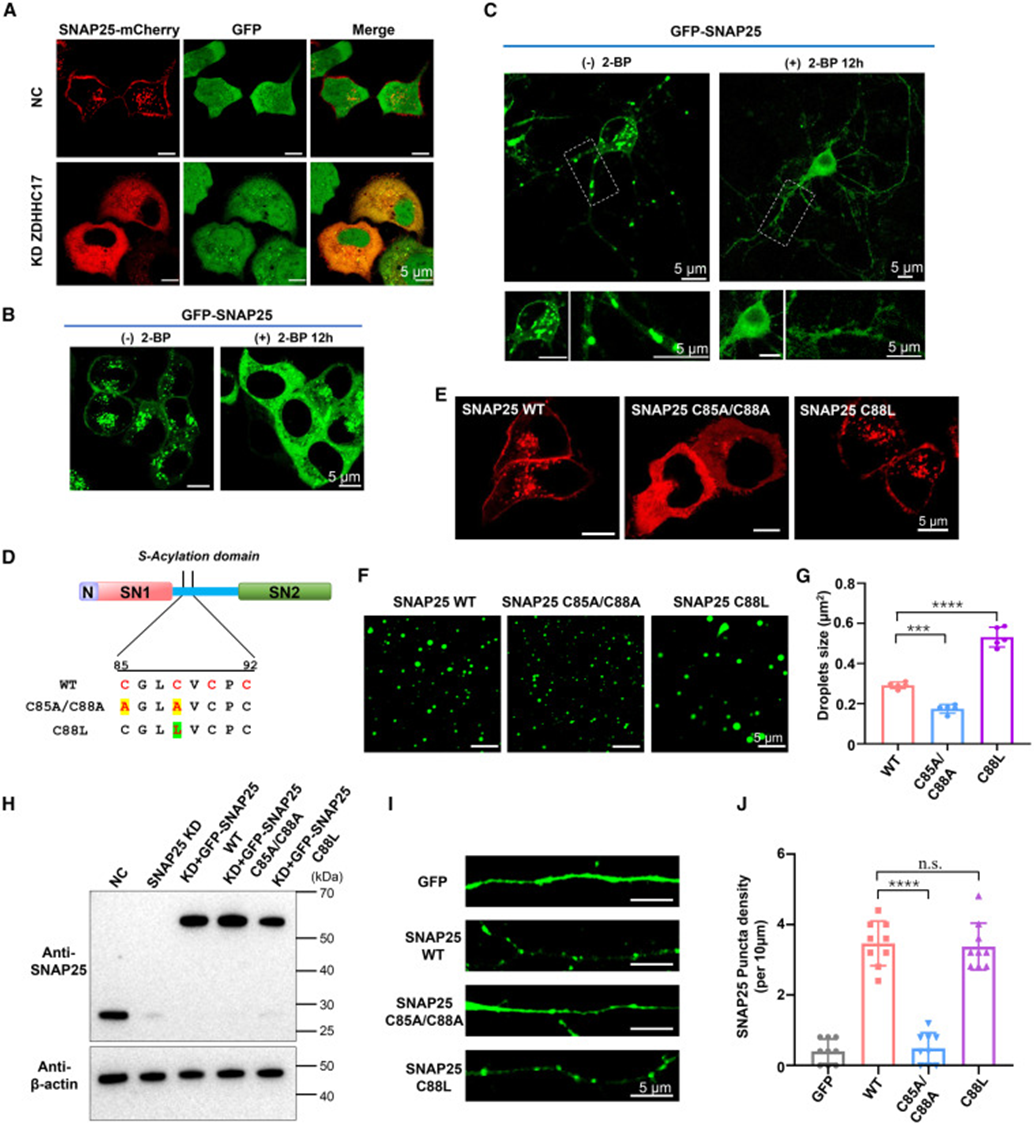
3. Palmitoylation modification bidirectionally regulates SNAP25 phase separation and membrane anchoring
Cysteines in the linker region are modification sites for palmitoyltransferase ZDHHC17. The study verified modification function through two approaches: cellular and in vitro mutation experiments. At the cellular level, knockdown of ZDHHC17 or addition of palmitoylation inhibitor 2-BP significantly reduced SNAP25 punctate condensates in HeLa and hippocampal neurons, with proteins detaching from cell membranes and distributing uniformly in the cytoplasm, indicating that palmitoylation deficiency directly disrupts condensate formation and membrane localization. The researchers constructed two types of site-directed mutations: C85A/C88A double mutation to eliminate cysteine modification sites, and C88L single mutation to simulate the hydrophobic effect of palmitoylation. In the in vitro phase separation system, double mutation showed significantly reduced droplet size and quantity, while the C88L mutation (simulating modification) produced larger and denser droplets. Supported lipid bilayer (SLB) membrane reconstitution experiments reproduced this trend—only proteins with palmitoylation sites could form high-density protein clusters on the membrane surface, and mutant proteins showed significantly weakened membrane aggregation ability. Primary neuron imaging also matched in vitro results: C88L mutation retained abundant on-membrane fluorescent spots, while double mutation showed diffuse distribution. Thus, it is confirmed that palmitoylation enhances linker region hydrophobicity, promoting SNAP25 liquid-phase condensation on one hand and mediating condensate anchoring on the synaptic cell membrane on the other.
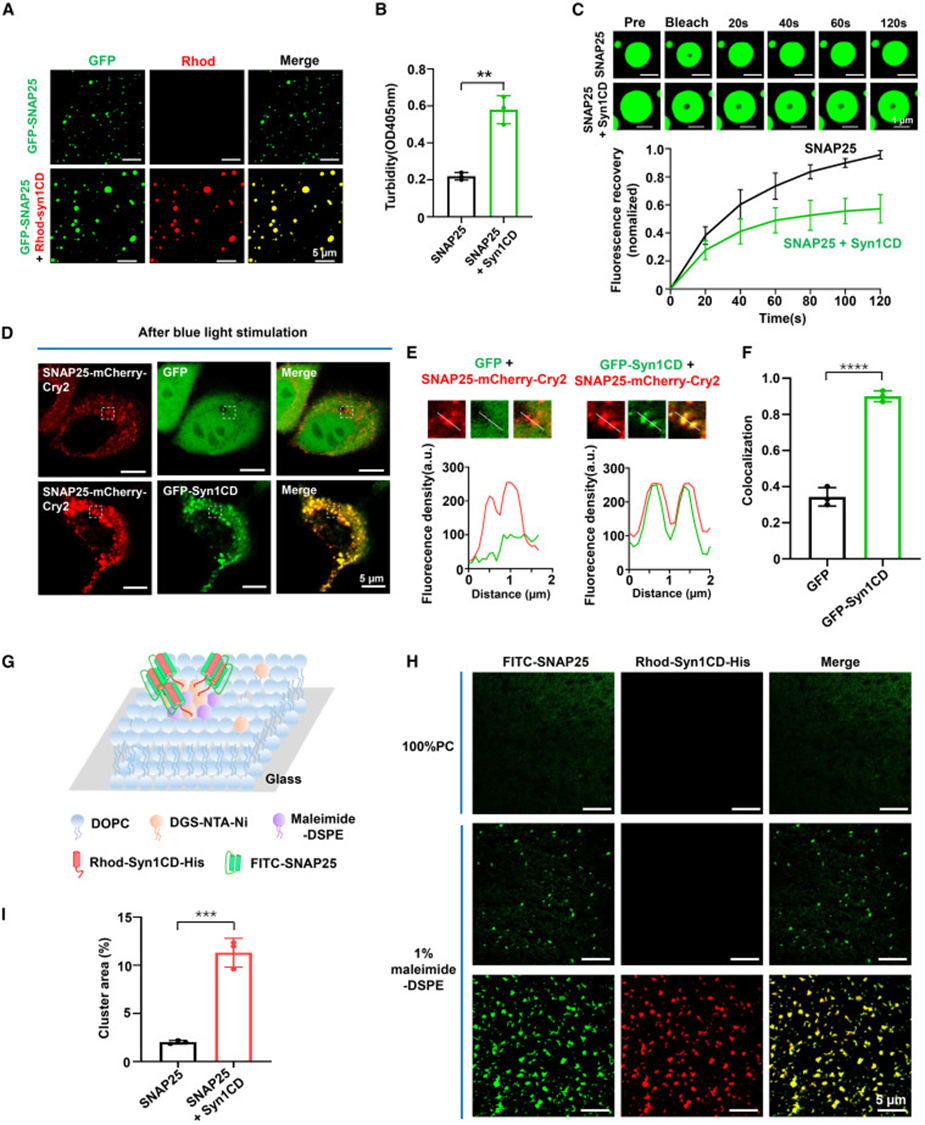
4. Syntaxin-1 co-condenses with SNAP25 and enhances phase separation efficiency
After mixing equal amounts of Syntaxin-1 cytoplasmic domain with SNAP25, dual-color fluorescence imaging showed complete co-localization of both proteins within the same droplet, with significantly increased droplet volume. Turbidity quantification values increased synchronously, FRAP experiments showed decreased protein mobility within co-condensates, and circular dichroism demonstrated that originally disordered SNAP25 converted to α-helical conformation after binding Syntaxin-1, indicating that their interaction induces protein structural folding. Experiments with gradient addition of Syntaxin-1 showed a dose-dependent effect—the proportion of SNAP25 in protein precipitation continuously increased with Syntaxin-1 concentration. In the OptoDroplet light-controlled co-expression system, Syntaxin-1 fluorescence completely infiltrated SNAP25 photo-induced condensation granules, and Pearson correlation coefficient confirmed their high co-localization. Artificial supported lipid bilayer system simulated cell membrane environment: after Syntaxin-1 was immobilized on the membrane, SNAP25 aggregated in large quantities to form larger co-condensation plaques. Together, both solution and membrane environments demonstrate that Syntaxin-1 can undergo co-phase separation with SNAP25 while amplifying condensation scale.
5. SNAP25 condensates recruit all SNARE components to form ternary co-condensates
In in vitro three-color fluorescence labeling experiments, both Syntaxin-1 and VAMP2 proteins were evenly distributed inside SNAP25 droplets. Sedimentation experiments recovered condensation precipitates containing all three SNARE proteins simultaneously, confirming that condensates possess the ability to enrich both partner proteins. When HeLa cells co-expressed three fluorescently labeled SNARE proteins, giant fused condensates formed within cells. FRAP results showed significantly reduced protein mobility within ternary co-condensates, suggesting extensive interactions between proteins. In vitro equimolar mixing of the three proteins directly formed large-sized co-condensation droplets. Supporting in vitro SNARE assembly experiments confirmed that functional SNARE complex production significantly increased under phase separation conditions. This clearly demonstrates that SNAP25 droplets serve as molecular scaffolds, simultaneously capturing both types of companion SNARE proteins to provide a local high-concentration microenvironment for four-helix fusion complex assembly.
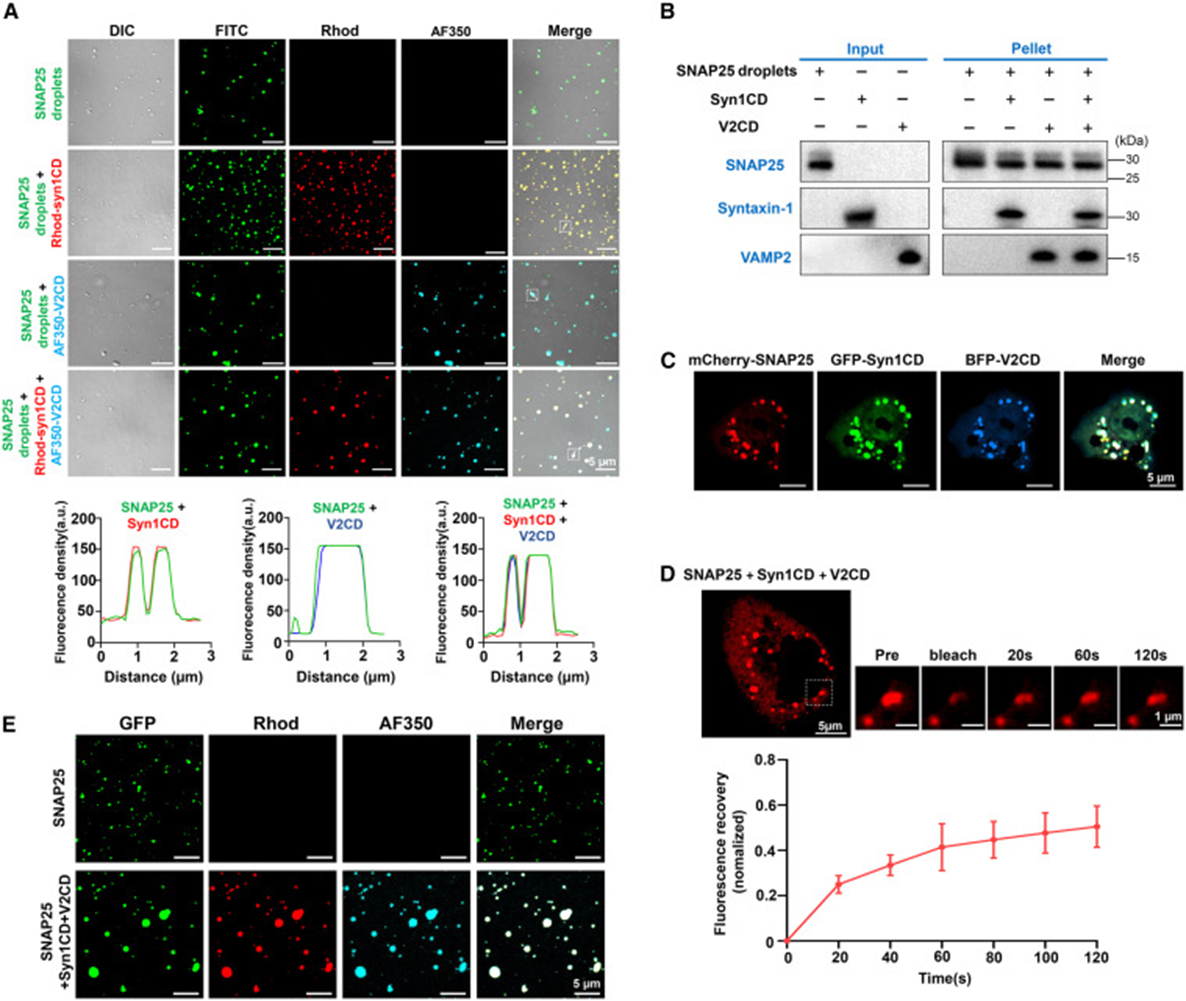
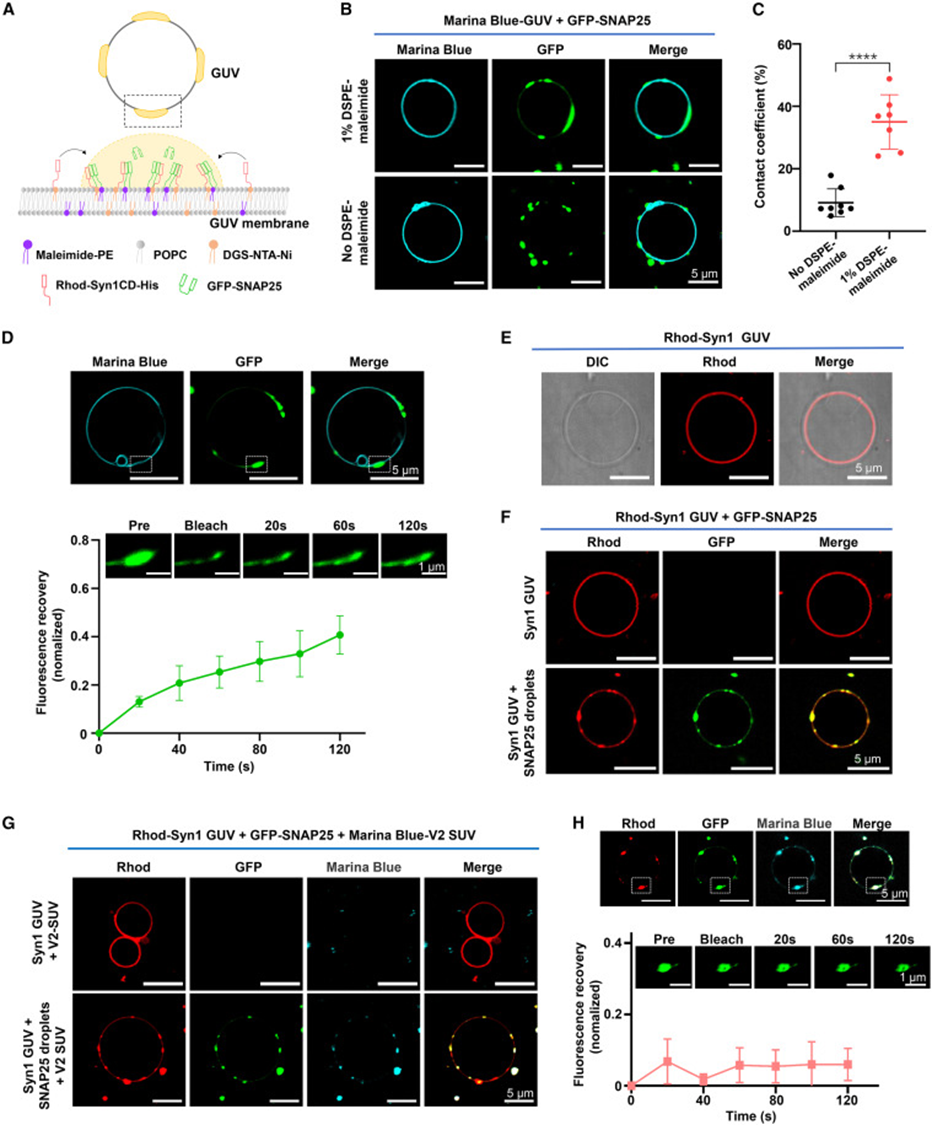
6. SNAP25 condensates adsorb to cell membranes and mediate synaptic vesicle anchoring
The study used GUV giant vesicles to simulate presynaptic cell membranes. Vesicles with maleimide lipids could bind SNAP25, and fluorescence imaging clearly observed SNAP condensates attaching to and infiltrating the vesicle surface. Removal of maleimide membrane-binding sites prevented droplets from adsorbing to the membrane structure. When Syntaxin-1 was pre-anchored on GUV membranes, adding SNAP25 formed large co-condensation regions on the membrane surface. Subsequent addition of VAMP-labeled small vesicles (SUVs) resulted in all vesicles concentrating within the membrane SNARE co-condensation plaques. Without SNAP25, vesicles could not anchor to the membrane surface. FRAP detection on membrane-bound ternary condensates showed significantly reduced protein mobility, confirming that multiple interactions between the membrane and three types of SNAREs stabilize the vesicle tethering state. This completely reproduces the physiological process: membrane-anchored SNAP25 condensates build a fusion platform that captures VAMP2-carrying synaptic vesicles, completing the vesicle docking step.
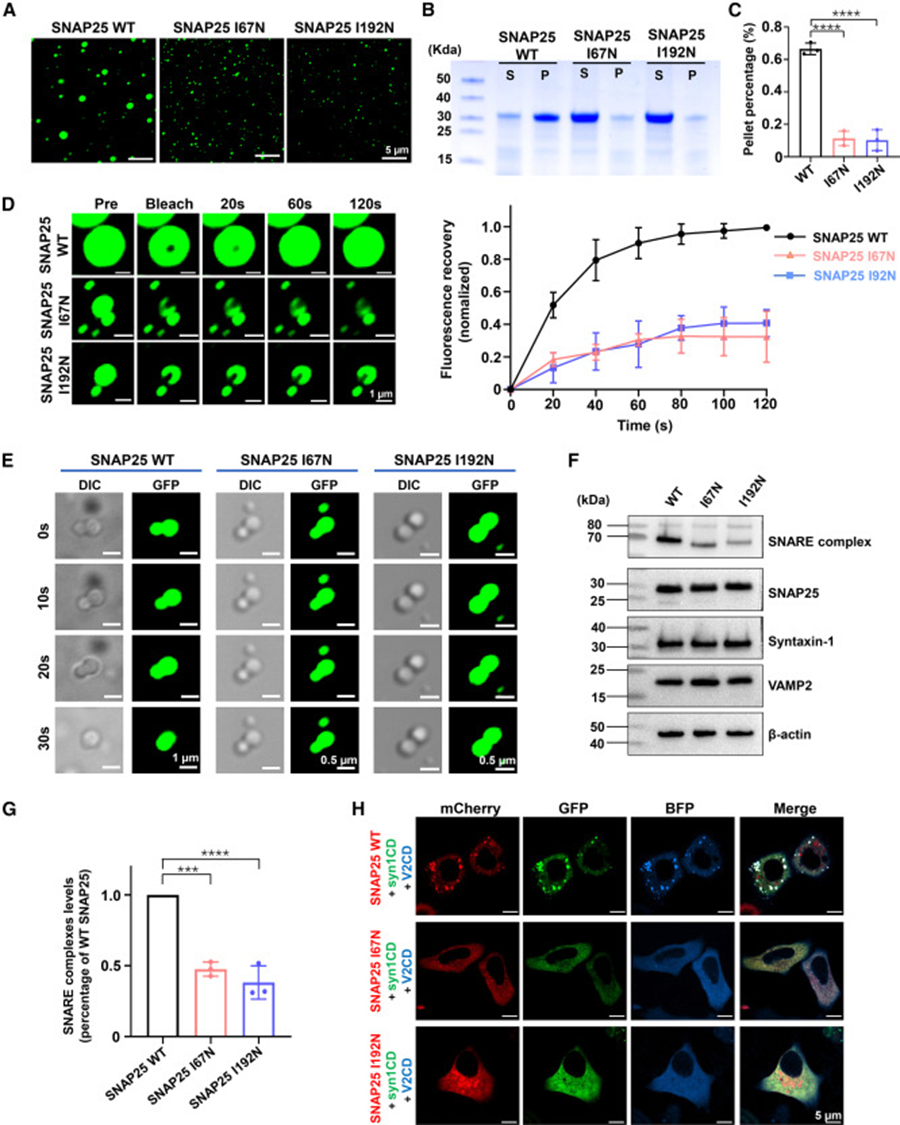
7. DEE pathogenic mutations disrupt phase separation and inhibit SNARE complex assembly
Two clinical DEE pathogenic mutations (I67N, I192N) were selected for functional verification. Under identical in vitro conditions, mutant proteins only formed tiny, sparse droplets. Sedimentation experiments showed significantly reduced protein entering condensation precipitates. FRAP curves demonstrated hindered molecular movement within mutant condensates, and time-lapse droplet fusion imaging showed that mutant small droplets could hardly fuse and grow, indicating that mutations reduce protein mobility and block droplet fusion maturation. At the cellular level, co-expression of three SNARE proteins showed abundant SDS-resistant functional SNARE complexes in wild-type groups, while both mutant groups showed significantly reduced complex content. In three-color fluorescence co-transfected cells, mutant SNAP25 failed to form large ternary co-condensates with Syntaxin-1 and VAMP2. These results establish a molecular pathological pathway: pathogenic mutations impair SNAP25 liquid-liquid phase separation ability, disrupt the ternary SNARE co-condensation platform, and ultimately inhibit membrane fusion machinery assembly, explaining the molecular underlying mechanism of impaired synaptic release in DEE patients.
Summary
This study systematically investigated SNAP25, the core protein of synaptic vesicle fusion, completely revealing the palmitoylation-modulated SNAP25 liquid-liquid phase separation mechanism, clarifying its core physiological functions in SNARE complex assembly, synaptic vesicle anchoring, and neurotransmitter release, and linking it to the pathogenesis of developmental epileptic encephalopathy. The study first confirmed through in vitro biochemistry and live-cell imaging that SNAP25 can undergo liquid-liquid phase separation, with the intrinsically disordered linker region between the two SNARE motifs being the essential structure for phase separation. Palmitoylation modification of cysteines in the linker region enhances protein hydrophobicity, dual-promoting condensate formation and cell membrane anchoring. Syntaxin-1 co-condenses with SNAP25, further amplifying the phase separation effect and inducing SNAP25 conformational changes, building a t-SNARE-enriched scaffold on the membrane surface. SNAP25 condensates can simultaneously recruit Syntaxin-1 and VAMP2 to form ternary SNARE co-condensates, significantly improving the assembly efficiency of functional SNARE complexes. Artificial membrane reconstitution experiments proved that membrane-bound SNAP25 condensates can infiltrate the membrane interface and capture VAMP2-containing synaptic vesicles, explaining the molecular basis of millisecond-scale synchronous neurotransmitter release in neurons dependent on local SNARE protein enrichment. The study further discovered that two DEE pathogenic mutations (I67N, I192N) disrupt SNAP25 condensate mobility, block droplet fusion, and inhibit ternary co-condensate and functional SNARE complex formation, establishing a link between phase separation defects and neurodevelopmental diseases at the molecular level. The article objectively notes current limitations: the regulatory effects of key regulatory proteins such as Munc18 and Munc13 on SNAP25 condensates have not been resolved, and in vivo animal experiments are lacking to corroborate physiological functions. Overall, the study fills the gap in the field of SNARE core protein phase separation regulating synaptic exocytosis, proposes a new paradigm of membrane-less condensates regulating rapid neuronal communication, and expands the understanding of presynaptic molecular arrangement and neurotransmitter release mechanisms.
References
Zhu M, Jin Y, Liu J, Kong X, Liu Y, Yu H. SNAP25 undergoes phase separation to facilitate the assembly of the synaptic vesicle fusion machinery. Cell Rep. 2026 Feb 24;45(2):116983. doi: 10.1016/j.celrep.2026.116983. Epub 2026 Feb 10. PMID: 41678336.